Department of Physiology, Development and Neuroscience
My research on the ears of non-mammalian vertebrates
The middle ear structures of birds, reptiles and frogs differ from those of mammals in that the tympanic membrane is coupled to the stapes, the only ear ossicle in these animals, by means of a cartilaginous structure called the extrastapes (the stapes and extrastapes of these animals are sometimes referred to as the "columella" and "extracolumella" respectively). Fishes lack a tympanic middle ear, but some species have Weberian ossicles to couple swimbladder vibrations to the sensory structures of the inner ear. Find out more by following the links below:
My research has largely concentrated on frog ears:
- My latest collaboration, with Dr. Sandra Goutte and an international team, has been to look at the hearing of pumpkin toadlets, some of the smallest frogs in the world (Fig. 1). Using a variety of techniques, we have found that these animals cannot actually hear their own calls (Goutte et al., 2017). Wondering how else they might communicate, we discovered that the bones just under their skin fluoresce when illuminated with UV light (Goutte et al., 2019). We cannot yet say for sure whether these tiny toadlets can see the patterns so revealed though.
- Working with Prof. Pim Van Dijk and colleagues, I have created 3D models of the inner ears of several frogs. We discovered that the classic illustration of the frog ear created by E.G. Wever, which has been used in textbooks, lectures and articles for many years, is inaccurate in several respects. See Mason et al. (2015) for more.
- I have investigated ear function in the aquatic frog Xenopus laevis, best-known as an animal model in developmental biology studies. This strange frog lacks a tympanic membrane but instead has a cartilaginous tympanic disc, formed as an expansion of the extrastapes. The ear of this animal, like that of the bullfrog, is sexually dimorphic, with male Xenopus having much larger tympanic discs than females. A rocking movement of the stapes is found in the Xenopus ear, as in the bullfrog, but in Xenopus the extrastapes (tympanic disc) is more rigidly coupled to the stapes and the resulting lever ratio is much smaller (Mason et al., 2009). This probably represents an adaptation to improve hearing underwater.
- Using a laser interferometer to make precise measurements of nanometer-scale movements, Prof. Peter Narins (of UCLA) and I were able to establish exactly how the middle ear apparatus vibrates in the bullfrog. We found that there is some flexibility between stapes and extrastapes (Fig. 2), and that a cartilaginous band called the ascending process of the extrastapes provides pivotal support. Although text-book illustrations tend to represent the stapes and extrastapes of frogs as simple pistons, the extrastapes of frogs actually works, in effect, as a second ossicle (Mason & Narins, 2002a)! Within the caudal half of the oval window in many frogs and salamanders is a second otic element called the operculum. Although long believed to be part of a separate pathway involved in seismic sensitivity, our laser interferometric work suggests that in bullfrogs the operculum is actually coupled to the stapes footplate: it moves in response to airborne sound (Mason & Narins, 2002b). The operculum may confer protection against quasi-static pressure changes associated with breathing and perhaps vocalization. Flexibility within the ossicular apparatus may turn out to be universal among terrestrial vertebrates, perhaps because of the advantages conferred with respect to pressure buffering. This is an intriguing avenue of research which I am continuing to explore (see Mason & Farr, 2013).
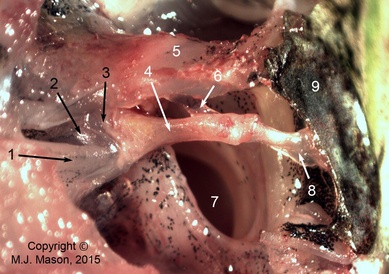
The middle ear apparatus of a frog
Photomicrograph of the right middle ear apparatus of a ranid frog, seen from dorsally, caudally and slightly laterally. 1: m. opercularis; 2: operculum; 3: stapes footplate; 4: stapes shaft (pars media); 5: parotic crest; 6: ascending process of extrastapes; 7:Eustachian tube; 8: extrastapes; 9: tympanic membrane.
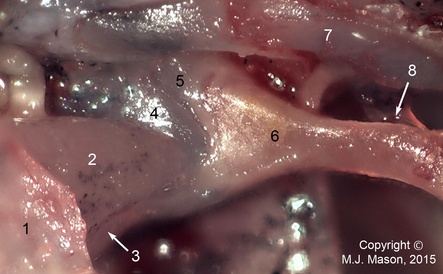
Close-up of the stapes footplate and otic operculum. 1: suprascapular cartilage; 2: m. opercularis; 3: m. levator scapulae superior aponeurosis; 4: operculum; 5: stapes footplate (pars interna); 6: bony stapes (pars media); 7: parotic crest; 8: ascending process of extrastapes.
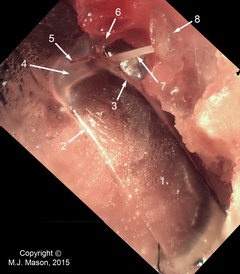
The middle ear apparatus of a bird
Photomicrograph of the right middle ear apparatus of a pheasant, internal view looking caudally. The inner ear and surrounding bone have been removed to reveal the stapes footplate. 1: tympanic membrane; 2: middle drum-tubal ligament; 3: infrastapedial process of extrastapes; 4: extrastapedial process of extrastapes; 5: Platner's ligament (cut); 6: suprastapedial process of extrastapes; 7: bony stapes shaft; 8: stapes footplate.
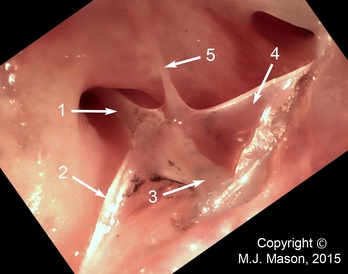
The left middle ear apparatus seen from externally, the tympanic membrane having been removed. 1: bony stapes shaft; 2: infrastapedial process of extrastapes; 3: extrastapedial process of extrastapes; 4: suprastapedial process of extrastapes; 5: Platner's ligament.
The Weberian apparatus of the zebrafish, Danio rerio
Rebecca Jordan (2011)
The Weberian apparatus is a specialized auditory structure of the axial skeleton of otophysan fish (series Otophysi, within the superorder Ostariophysi). It comprises four bony elements or Weberian ossicles, derived from parts of the anterior-most vertebrae, which via their ligamentous connections form a chain linking the swimbladder to the inner ear. From posterior to anterior, the Weberian ossicles are the tripus, intercalarium, scaphium and claustrum. The tripus is connected via ligaments to the anterior portion of the swimbladder, while the anterior elements are coupled to the inner ear fluid space (the atrium of the sinus impar). The Weberian ossicles mechanically couple vibrations of the swimbladder induced by sound to the inner ear fluid, allowing sound to be transduced into electrical signals by the sensory cells there. The function of the Weberian ossicles is therefore in some respects analogous to that of the middle ear ossicles of terrestrial vertebrates, although these structures are completely unrelated.
There is considerable variability in the structure of the Weberian apparatus among the numerous species of otophysan fish (Ladich & Popper, 2004). We investigated the morphology of the zebrafish Weberian apparatus through micro-CT scanning and alizarin red staining: some of our results are shown as Figs. 1-3 below. In zebrafish, the tripus is the largest Weberian ossicle, connected via the very delicate intercalarium to the scaphium. The claustrum, located dorsal to the scaphium (Fig. 2), is described as being fused to the first vertebra and thus immobile in zebrafish, functioning to support the wall of the sinus impar atrium (Bang et al., 2001). Development of the Weberian apparatus as zebrafishes grow coincides with an expansion of maximum frequency detection from 200 to 4000 Hz, suggesting that these structures have a role in the detection of higher sound frequencies (Higgs et al., 2002). It is becoming clear as research progresses that the ears of fishes are much more specialized than previously believed.
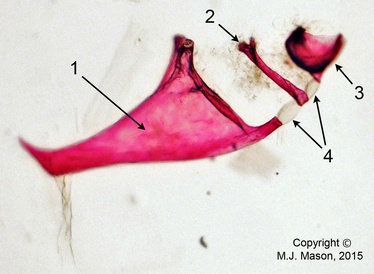
Fig. 3. Photomicrograph (taken at 40x magnification) of three of the four Weberian ossicles dissected from an adult male zebrafish. The tripus (1), intercalarium (2) and part of the scaphium (3) are still attached in their chain via inter-ossicular ligaments (4). The fish had been fixed with PFA and stained using the alizarin red staining process, rendering the flesh transparent and calcified matrix (such as bone) pink.
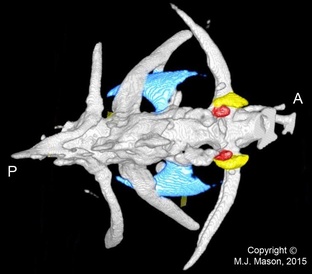
Fig. 1. Micro-CT image of the most anterior vertebrae of an adult male zebrafish, dorsal view. The tripus is coloured blue, the scaphium yellow and the claustrum red. A = anterior; P = posterior. The intercalarium, which is located between the scaphium and tripus, is not visible in this reconstruction, presumably due to scan resolution limitations.
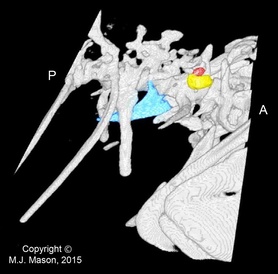
Fig. 2. Right lateral view of the same micro-CT reconstruction.
References
Bang, P.I., Sewell, W.F. & Malicki, J.J. (2001) Morphology and cell type heterogeneities of the inner ear epithelia in adult and juvenile zebrafish (Danio rerio). Journal of Comparative Neurology 438: 173-90.
Higgs, D.M., Rollo, A.K., Souza, M.J. & Popper, A.N. (2003) Development of form and function in peripheral auditory structures of the zebrafish (Danio rerio). Journal of the Acoustical Society of America 113: 1145-54.
Ladich, F. & Popper, A.N. (2004) Parallel evolution in fish hearing organs. In: Evolution of the Vertebrate Auditory System (eds Manley, G.A., Popper, A.N. & Fay, R.R.), pp. 95-127. New York: Springer.
This work represents the results of a short undergraduate project performed in the Mason and Fleming laboratories by Rebecca Jordan in 2011.
Please contact Dr. Matthew Mason for more information about this as yet unpublished work.